Research Projects
Doctoral Research
Yale School of the Environment
Advisors
Mark A. Bradford, Ph.D. & Oswald J. Schmitz, Ph.D.
Soil Inoculation Experiment Preliminary Findings
Presented at the British Ecological Society Meeting 16 December 2025
Background
One challenge in managed forests can be regenerating commercially and biodiverse high value species, many of which are mid-to-late successional species such as oak and hickory (Hanewinkel et al. 2013). Economically, foresters and land owners depend on these high value species (Hannah 1987), which also have high ecological value. In the Northeastern US, red oak is in decline and faces regeneration challenges (Oliver et al. 2005, Payne and Peet 2023). The failure to regenerate red oak and hickory is often attributed to site conditions, shade tolerance, or modern fire suppression (Payne and Peet 2023) but no one has examined how below-ground biotic interactions may be influencing successes. These species associate with ectomycorrhizal fungi (ECM), which expands their rooting network and function, allowing the trees to access nutrients in organic as well inorganic forms. In managed forests, ECM connectivity is sometimes limited, which may be impeding oak and hickory regeneration (Cortese and Horton 2023). One solution to redress limitation of mycorrhizal inoculum has been to simply add the mycorrhizal fungal inocula to regenerating species.
The use of inoculants in other ecosystem types has led to commercial mycorrhizal inoculants being marketed to forest managers in temperate forests (i.e., Mycorrhizal Applications) but their effectiveness for establishing desired tree species appears untested at commercial scales such as at the stand and forest level. Furthermore, research in other systems has suggested that transplanting a thin layer of soil, as an alternative means to inoculate beneficial organisms, may be more effective than commercial inoculum to improve regeneration success of high value species, by also establishing native species opposed to foreign ones (Li et al. 2024).
For instance, we know from grassland systems that adding a local soil from a late successional stage grassland to a regenerating one can shift the community composition toward later successional stages (Wubs et al. 2019) and that adding arbuscular mycorrhizal fungi in grasslands often improves late successional species success (Allen and Allen 1984). Further, tests comparing commercially available inoculum and local soil inoculum have demonstrated that local soil inoculum can improve late successional species success, but commercial inoculum does not (Rowe et al. 2007, Emam 2016). However, the success of commercial inoculum versus a local soil inoculum and their impact on tree species success has not been addressed in eastern temperate forests, despite an interest by practitioners in belowground management.
In addition, the introduction of soil fauna through whole soil inoculation may mediate the effect of inocula through nutrient deposition from soil animal bodies and frass (Hunt et al. 1987), by grazing on mycorrhizal hyphal networks (Crowther et al. 2012), and indirectly through trophic cascades (McCary and Schmitz 2021, Lienau et al. 2024), thus altering the potential impact of inocula on plant growth and providing longer term diversity and stability in the system. Direct comparisons of commercial vs. local whole soil inoculant are lacking in regenerating forests and must be tested for their effectiveness on high value tree species regeneration, success or failure for establishing a limited belowground community, and tests for the ecosystem response. Thus, this experiment seeks to answer the following questions:
Questions
(1) Does late successional soil inoculation improve aboveground seedling growth in a shelterwood stand?
(2) Does soil inoculation successfully establish a belowground soil community similar to the late successional forest?
(3) Does soil inoculation alter belowground soil processes such as decomposition rates, nutrient cycling, soil pH, soil moisture, or water holding capacity?
Methods
We initiated an experiment at Yale-Myers Forest (YMF) in CT, USA at the beginning of the growing season in 2024 in a recently harvested shelterwood forest stand, (Stand 5074) which is managed for high value hardwoods such as red oak and hickory species. We established 20 blocks, each clustered with four, 2 x 2-m treatment plots. Plots were spaced > 5 m apart resulting in each of the 20 larger blocks of ~ 11 x 11 m. We spaced each block 20 m apart. The resulting experiment area covered 135 x 104 m (1.3 ha). Given Stand 5074 is covered in a large amount of slash, we also haphazardly selected plot locations and cleared all the slash > 0.5 m long within the 2 x 2 m treatment plot areas (Fig. 1) (n = 20 blocks x 4 plots = 80).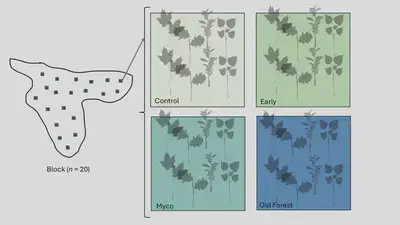
To test the effectiveness of commercial inoculum versus late successional whole soil transplants, we compared treatments with two controls. These were two “treatment” plots and two “control” plots partitioned as follows: i) local early succession soil inoculum control, ii) a control where nothing is added, iii) local late successional soil inoculum, and iv) a common commercial inoculum.
We sourced the early successional soil from the experimental stand, by applying an 80 x 80 x 10 cm volume of soil and litter sourced from a location directly next to each plot. The late successional soil was sourced from a stand nearby (Stand 2001). The source location met the following criteria i) no invasive species (i.e., earthworms, EAB) and ii) mix of deciduous species from our seedling mixture, iii) healthy stand with trees > 100 years. From 20 of the randomly assigned unique locations in the late successional reference stand, we took an 80 x 80 x 10 cm volume of soil and litter and placed them into 5-gallon buckets and transferred them to the experiment site. These 20 soil cubes were directly spread on the treatment plots the same day (Fig. 2).
We further tracked the belowground responses to understand the mechanisms of interaction between the soil and plant communities. We expected that transplanting the soil community will change the fundamental soil function via changes in the soil community. We measured overall microbial biomass, soil organic carbon and nitrogen, inorganic nitrogen, carbon mineralization, pH, soil moisture, and water holding capacity. We took initial baseline soil samples before treatment in each plot and repeated soil sample collection each spring and will through 2026, and will take a final soil sample in the fall of 2026 to track changes over time and capture natural variation among plot locations.
Seedling growth was calculated by subtracting the initial seedling stem height at planting in May from the final stem height in October of the second year. We then compared seedling response to treatment using linear models from lmer in R Studio with plot ID nested within Block ID as a random effect (Bates et al. 2015). We then extracted coefficients, standard error, and P-values in original units from lmerTest, considering P values < 0.05 significant (Kuznetsova et al. 2017). We used the same model structure for all soil variables with block as a random effect, since the data was at the plot level.
Preliminary Findings
Aboveground
Tree seedling survival during the first and second growing season was even across treatments (71-3 percent), although slightly lower in the control at 61 percent (Fig. 3a). Seedling survival was lowest for sugar maples (41 percent) (Acer saccharum; ACSA; Fig 3b), 77 percent for black birch (Betula lenta; BELE), 81 percent for black cherry (Prunus serotina; PRSE), and high at 91 percent for red oak (Quercus rubra; QURU). 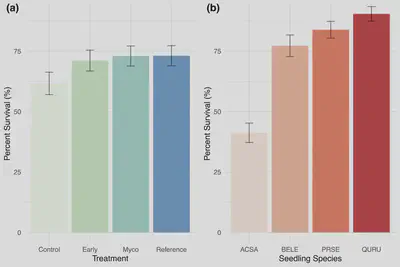
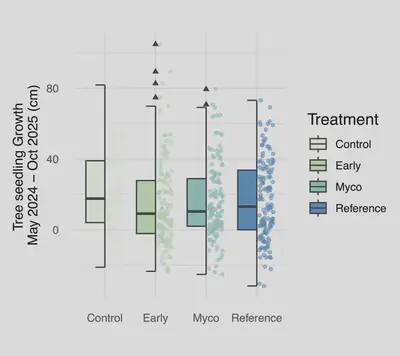
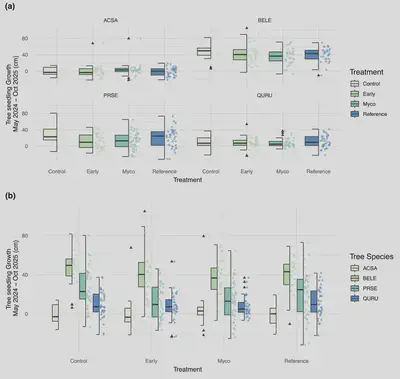
Belowground
Belowground soil function responded to treatment and controls after two growing seasons (Fig. 6). Soil pH was elevated under the early soil inoculation treatments after one year (Estimate: 0.14550 ± 0.05964, P = 0.0178), while SIR (Estimate: 2.8342 ± 1.2648, P = 0.029), carbon mineralization (Estimate: 96.27± 16.61, P = 3.11e-07), and water holding capacity (Estimate: 0.19446± 0.05341, P = 0.000587) were all greater under the reference whole soil inoculation treatment.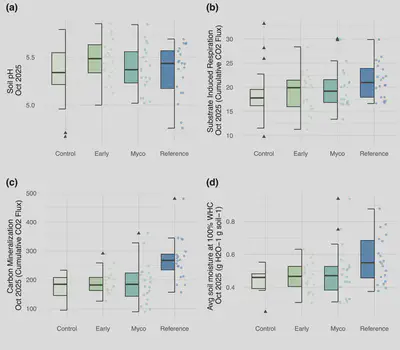
References
Allen, E. B., and M. F. Allen. 1984. Competition between plants of different successional stages: mycorrhizae as regulators. Canadian Journal of Botany 62:2625–2629.
Bates, D., M. Mächler, B. Bolker, and S. Walker. 2015. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software 67:1–48.
Borden, C. G., M. C. Duguid, and M. S. Ashton. 2021. The legacy of fire: long-term changes to the forest understory from periodic burns in a New England oak-hickory forest. Fire Ecology 17:1–17.
Contos, P., N. P. Murphy, and H. Gibb. 2023. Whole-of-community invertebrate rewilding: Leaf litter transplants rapidly increase beetle diversity during restoration. Ecological Applications 33:e2779.
Contos, P., N. P. Murphy, Z. J. Kayll, T. Morgan, J. J. Vido, O. Decker, and H. Gibb. 2024. Rewilding soil and litter invertebrates and fungi increases decomposition rates and alters detritivore communities. ECOLOGY AND EVOLUTION 14:e11128.
Contos, P., J. L. Wood, N. P. Murphy, and H. Gibb. 2021. Rewilding with invertebrates and microbes to restore ecosystems: Present trends and future directions. Ecology and Evolution 11:7187–7200.
Cortese, A. M., and T. R. Horton. 2023. Islands in the shade: scattered ectomycorrhizal trees influence soil inoculum and heterospecific seedling response in a northeastern secondary forest. Mycorrhiza 33:33–44.
Crowther, T. W., L. Boddy, and T. Hefin Jones. 2012. Functional and ecological consequences of saprotrophic fungus–grazer interactions. The ISME Journal 6:1992–2001.
Emam, T. 2016. Local soil, but not commercial AMF inoculum, increases native and non-native grass growth at a mine restoration site. Restoration Ecology 24:35–44.
Frouz, J., A. Roubíčková, P. Heděnec, and K. Tajovský. 2015. Do soil fauna really hasten litter decomposition? A meta-analysis of enclosure studies. European Journal of Soil Biology 68:18–24.
Hanewinkel, M., D. A. Cullmann, M.-J. Schelhaas, G.-J. Nabuurs, and N. E. Zimmermann. 2013. Climate change may cause severe loss in the economic value of European forest land. Nature Climate Change 3:203–207.
Hannah, P. R. 1987. Regeneration Methods for Oaks. Northern Journal of Applied Forestry Volume 4:Pages 97-101. Hunt, H. W., D. C. Coleman, E. R. Ingham, R. E. Ingham, E. T. Elliott, J. C. Moore, S. L. Rose, C. P. P. Reid, and C. R. Morley. 1987. The detrital food web in a shortgrass prairie. Biology and Fertility of Soils 3:57–68.
Ladau, J., A. K. Fahimipour, M. E. Newcomer, J. B. Brown, G. J. Vora, M. K. Melby, and J. A. Maresca. 2023. Microbial invasions and inoculants: a call to action.
Li, C., X. Chen, Z. Jia, L. Zhai, B. Zhang, U. Grüters, S. Ma, J. Qian, X. Liu, J. Zhang, and C. Müller. 2024. Meta-analysis reveals the effects of microbial inoculants on the biomass and diversity of soil microbial communities. Nature Ecology & Evolution 8:1270–1284.
Lienau, J. R., M. C. Duguid, and O. J. Schmitz. 2024. Ground beetle trophic interactions alter available nitrogen in forest soil. Oikos:e10638.
McCary, M. A., and O. J. Schmitz. 2021. Invertebrate functional traits and terrestrial nutrient cycling: Insights from a global meta-analysis. Journal of Animal Ecology 90:1714–1726.
Oliver, C. D., E. C. Burkhardt, and D. A. Skojac. 2005. The increasing scarcity of red oaks in Mississippi River floodplain forests: Influence of the residual overstory. Forest Ecology and Management 210:393–414.
Payne, C. J., and R. K. Peet. 2023. Revisiting the model system for forest succession: Eighty years of resampling Piedmont forests reveals need for an improved suite of indicators of successional change. Ecological Indicators 154:110679.
Rowe, H. I., C. S. Brown, and V. P. Claassen. 2007. Comparisons of Mycorrhizal Responsiveness with Field Soil and Commercial Inoculum for Six Native Montane Species and Bromus tectorum. Restoration Ecology 15:44–52.
Werden, L., R. Cole, K. Schönhofer, K. Holl, R. Zahawi, C. Averill, D. Schweizer, J. Calvo-Alvarado, D. Hamilton, F. Joyce, M. San-José, F. Hofhansl, L. Briggs, D. Rodríguez, J. Tingle, F. Chiriboga, E. Broadbent, G. Quirós-Cedeño, and T. Crowther. 2024. Assessing innovations for upscaling forest landscape restoration. ONE EARTH 7:1515–1528.
Wubs, E. R. J., W. H. van der Putten, M. Bosch, and T. M. Bezemer. 2016. Soil inoculation steers restoration of terrestrial ecosystems. Nature Plants 2:16107.
Wubs, E. R. J., W. H. van der Putten, S. R. Mortimer, G. W. Korthals, H. Duyts, R. Wagenaar, and T. M. Bezemer. 2019. Single introductions of soil biota and plants generate long-term legacies in soil and plant community assembly. Ecology Letters 22:1145–1151.
MESc Research
Yale School of the Environment
Advisors
Oswald J. Schmitz, Ph.D. & Marlyse C. Duguid, Ph.D.
The main goal of my thesis was to investigate the relationship between soil food web predators and nitrogen cycling in eastern temperate forests. The ground beetle family (Coleoptera: Carabidae) has some of the most abundant predators in forest soil food webs. Despite a long history of carabid study, their role in nitrogen cycling remains largely undescribed. As predator generalists, they likely control populations of ants, mites, millipedes, and other invertebrate decomposer groups that fragment plant litter and influence nitrogen cycling. I conducted two experiments at the Yale-Myers Forest in eastern Connecticut. My hope is to better understand how these trophic interactions in soil invertebrate groups are important for decomposition, nitrogen cycling, and ecosystem function.
Check out my research spotlight in The Forest School’s newsletter
Trophic Cascade Project
The leading question for the first chapter of my master’s work was How do functional differences in ground beetles contribute to nitrogen cycling in eastern temperate old and young forests? To address this question we set up a trophic cascade experiment using mesocosm cages (pictured below) with live ground beetles. We measured the response of available nitrogen over the 68-day experiment. We set up cages in both young and old forest plots. Our results support a growing body of literature discussing how predators influence nitrogen cycling. Here our results suggest available nitrogen increases with the addition of predators, at least on a short time scale. Stay tuned for the publication.
Nitrogen Tracer Experiment
To better understand how larger groups of soil animals interact with functional groups of ground beetles and contribute to nitrogen cycling, we set up an experiment using a stable isotope of nitrogen (15-N) to trace the flow of nitrogen from the soil litter layer to invertebrate groups and ground beetles collected in pitfall traps. The 15-N was applied through leaf litter, and due to a drought in the summer of 2022 and lack of decomposition, we didn’t detect a 15-N signal in the soil or invertebrate groups. We’re running the experiment for a second (and maybe a third!) year to see what happens. Stay tuned for the publication.
Undergrad Research
The Morton Arboretum, Center for Tree Science
Advisors
Meghan G. Midgley, Ph.D. & Robert W. Buchkowski, Ph.D.
I conducted research under the mentorship of Drs. Meghan Midgley and Robert Buchkowski in the Soil Ecology lab at The Morton Arboretum’s Center for Tree Science in Lise, Illinois from January 2020 – August 2021. We carried out two projects on how tree species influence soil properties and invertebrate food webs in temperate forests.
Summer 2021 - REEF
As a Research Experience Extension Fellow (REEF) I was able to return to the arboretum for a second year so we could carry out an experiment in the arboretum monoculture forestry plots on how tree traits predict the abundance and diversity of soil macroinvertebrates across tree species. We set up cover boards, a quick way to survey macroinvertebrates, in 18 forestry plots and are tying tree and soil properties to the invertebrate community composition. Our research suggests tree and soil traits predict macroinvertebrate communities! Stay tuned for the publication.
Summer 2020 - REU
My covid summer research with the arboretum as an NSF Research Experiences for Undergraduates (REU) student meant an opportunity for me to learn RStudio. We shifted our original project to use data that was publically available from NSF’s National Ecological Observatory Network (NEON) archive of ground beetle data. Turns out, the abundance of evergreen trees really influences ground beetle communities! Stay tuned for the publication.
Thanks for reading!🪲